【摘要】 目的 探索骨髓间充质干细胞(bone mesenchymal stem cells,BMSCs)对子宫内膜癌Ishikawa细胞转录水平的影响。方法 用Lenti-EGFP慢病毒感染Ishikawa细胞,分别将表达增强型绿色荧光蛋白(enhanced green fluorescent protein,EGFP)的Ishikawa细胞单独培养、与BMSCs接触共培养;通过流式细胞术分离Ishikawa细胞,并进行转录组测序分析筛选差异表达的mRNA和miRNA,通过miRWalk预测差异miRNA靶基因,对两者的交集基因进行GO、KEGG、蛋白网络互作分析;采用qRT-PCR和Western blot验证测序结果的准确性。结果 标记EGFP的Ishikawa细胞与BMSCs共培养后,分离出的Ishikawa-EGFP阳性细胞占33.6%,阴性细胞占10.5%。测序后共筛选出5 928个差异表达mRNA和111个差异表达miRNA。蛋白网络互作分析显示交集基因表达的蛋白之间相互作用,核心节点包括CDKN1A、JAK1、COL1A1、VCAN等。miRNA-mRNA网络图显示CDKN1A与hsa-let-7e-5p存在潜在靶向关系。GO分析和KEGG分析结果显示,其交集基因参与细胞外基质结构、细胞黏附分子结合等,并主要富集在PI3K-Akt、黏着斑、MAPK、EGFR酪氨酸激酶抑制剂耐药等信号通路。qRT-PCR和Western blot结果显示,共培养后Ishikawa-EGFP细胞中CDKN1A表达上调,hsa-let-7e-5p表达下调。结论 BMSCs可能通过细胞间的相互作用调节肿瘤局部微环境并促进子宫内膜癌的发生发展。
【关键词】 子宫内膜癌;骨髓间充质干细胞;共培养;转录组测序
Volume14 Number3 June2022
01
—
材料与方法
1.1 主要材料与试剂
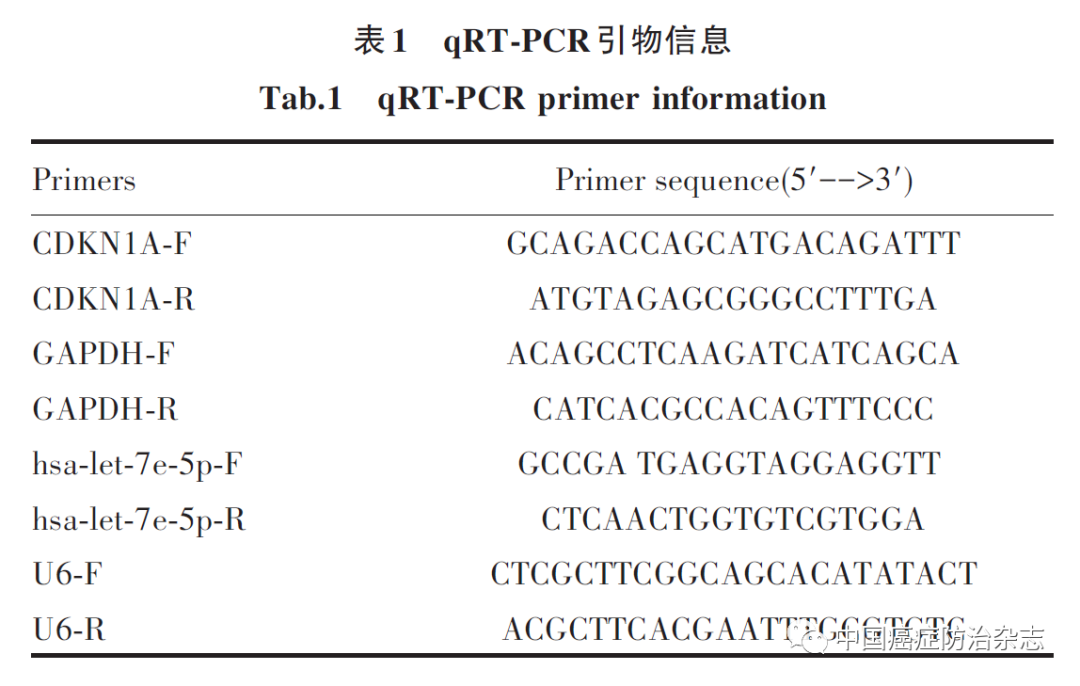
Ishikawa细胞购自北纳生物,人BMSCs、干细胞专用培养基购自上海赛百康生物科技有限公司,Lenti-EGFP慢病毒和Polybrene购自上海锐赛生物技术有限公司,高糖DMEM培养基购自江苏凯基生物技术股份有限公司,灭稻瘟素(blasticidin S,BSD)购自美国Millipore公司,10%胎牛血清(FBS)购自美国Gemini公司,0.25%胰酶、青霉素-链霉素溶液(P/S)、RIPA裂解液、BCA蛋白定量试剂盒、超敏ECL化学发光试剂盒购自上海碧云天公司,TRIzol总RNA抽提盒、反转录试剂盒购自..Takara公司,SYBR Green荧光定量试剂盒、PCR仪购自美国赛默飞公司,荧光激活细胞分选仪购自BD公司(BD AccuriTM C6),兔抗人ETS1、细胞周期蛋白依赖性激酶抑制剂1A(cyclin dependent kinase inhibitor 1A,CDKN1A)、GAPDH抗体、辣根过氧化物酶 (HRP)标记的二抗(兔抗)购自英国Abcam公司,qPCR引物由上海生工有限公司合成。
1.2 方法
第4天分别收集3组细胞,制备成单细胞悬液。通过BD AccuriTM C6在488 nm波长激发光下分离Ishikawa-EGFP+BMSCs组中EGFP阳性(EGFP+)和EGFP-细胞。空白组作为阴性对照样本调试电压,上样后调节细胞流速,维持在2 500 events/s左右,根据细胞大小和颗粒度选择Forward Scatter (FSC)和Side Scatter (SSC)范围,调整门的位置、大小和形状,调节荧光通道电压。取下对照样品管,上共培养组样品,开始采样。取有EGFP+细胞的采样管,10 000 r/min离心5 min,弃上清,PBS清洗1遍,再次离心后弃上清液。然后收集共培养后的Ishikawa-EGFP细胞,液氮速冻后于-80 ℃保存。将单独培养和共培养后的Ishikawa-EGFP细胞送上海美吉生物医药科技有限公司进行转录组测序分析。
1.3 统计学方法
采用SPSS 23.0及GraphPad Prism 8.0统计软件进行分析,计量资料以均数±标准差([x]±s)表示,两组间比较采用两独立样本t检验。双侧P<0.05表示差异有统计学意义。
02
—
结果
2.1 Ishikawa-EGFP细胞共培养后的增殖情况
采用荧光显微镜观察共培养组第1~4 天的绿色荧光细胞状态并计数,结果显示,EGFP标记的Ishikawa细胞数分别为94个、99个、145个、203个,表明共培养1~4 d的 Ishikawa-EGFP细胞保持增殖趋势,于第4天收集生长状态良好的细胞进行分选。见图1。
2.2 荧光激活细胞分选共培养后的Ishikawa-EGFP细胞
在BD AccuriTM C6分选仪上设门圈出空白组的Ishikawa细胞群,该群占细胞总数82.1%,见图2A;再以相应参数分选出Ishikawa-EGFP+BMSCs共培养组中带绿色荧光的Ishikawa-EGFP细胞,结果显示,Ishikawa-EGFP阴性细胞占细胞总数的10.5%,Ishikawa-EGFP阳性细胞占细胞总数的33.6%,见图2B。
2.3 筛选差异表达的mRNA和miRNA
原始测序数据按上述流程处理后,共筛选出5 928个差异表达mRNA,111个差异表达miRNA,且仅有65个miRNA 在miRWalk中可匹配到靶基因,其中表达下调16个,表达上调49个;共富集到2 701个靶基因,韦恩图显示其与差异表达mRNA有852个交集基因,其中548个表达上调,304个表达下调,见图3。PPI分析结果显示交集基因表达的蛋白之间相互作用(|Log2FC|≥2,P<0.05,蛋白互作分数=0.95),共有79个节点,68条边(即节点互作关系)。核心节点包括CDKN1A、JAK1、COL1A1、VCAN等,其中COL1A1与P4HA2、COL4A1、COL4A2之间存在互作关系(见图4A),且P4HA2、COL4A1、COL4A2表达上调(见图4B)。选取下调的16个差异表达miRNA及其预测到的交集基因,利用Cytoscape绘制miRNA-mRNA网络图,结果显示:hsa-let-7e-5p、hsa-let-7a-5p、hsa-let-7b-5p调控大量靶基因,其中hsa-let-7e-5p潜在靶基因包括CDKN1A,见图4B。
2.4 GO功能分析和KEGG富集分析结果
GO分析结果显示,上调的交集基因主要参与细胞外基质(extracellular matrix,ECM)结构、细胞膜结构、细胞黏附分子结合等,而下调交集基因则主要参与轴突发生、突触前活动区、生殖系统发育等,见图5;KEGG富集分析结果提示,上调差异基因主要富集在PI3K-Akt、黏着斑、MAPK、癌症中的miRNA、JAK-STAT、Ras等信号通路上,下调差异基因则主要富集在前列腺癌、EGFR酪氨酸激酶抑制剂耐药等通路,见图6。



![早啊,健康来了![2022.7.19]](http://img3.ii77.com/mmbiz_gif/OrSt8KRcTAwiayZcmhjSnB44R91cc3SgibqLdwK2tkS27mkGkvGia3R6WOuGlcPEgmbqtWYic6eBlXlZiaGwKGicZVFw/640?wx_fmt=gif&wxfrom=5&wx_lazy=1)